奇蹄目(きていもく、Perissodactyla)は、哺乳綱に分類される目。別名ウマ目。現生のウマ科、サイ科、バク科の他、多くの化石群が属す。草食性で、蹄を持ち、第3指(中指)に体重が乗る形で指が進化したグループであり、後肢の指は1本か3本の奇数である。
進化
奇蹄目は始新世から漸新世にかけて繁栄し、特に漸新世には有角亜目から陸上哺乳類史上最大級の種(パラケラテリウムまたはインドリコテリウム)が現れるなど、その繁栄の絶頂にあった。
しかし中新世以降は地球の寒冷化による環境の変化によって森林が減少し、草を食べることができなかった多くの種が絶滅し、さらに反芻亜目の進化に押されて衰退を始める。地質時代には240属と多様性を誇ったこのグループも、現在はわずかに3科6属20種しか生き残っていない。初期の奇蹄目は一般的な哺乳類の歯の数であったが、しだいに歯冠が高くなり、小臼歯が大臼歯になった。
草原を早く駆け抜けるのに適した分類群は尺骨と橈骨が、腓骨と脛骨が癒合し、脚を捻りにくくなり、天敵から逃げ切れるようになった。
起源
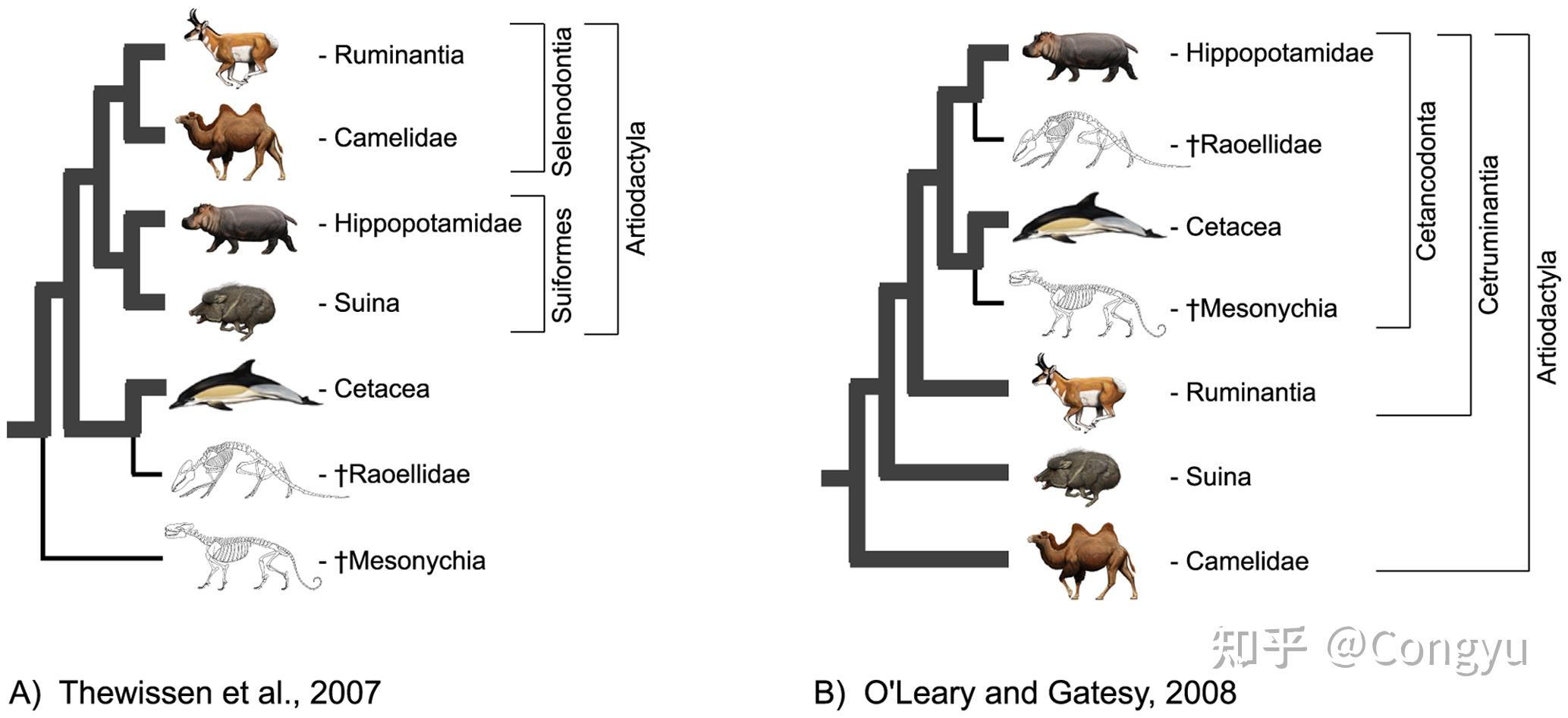
従来の説においては、奇蹄目と偶蹄目は約6000万年前に顆節目を祖として分岐、進化したとされてきた。
しかし2006年、分子生物学の見地から、奇蹄目は食肉目、翼手目などと近縁とされ、ローラシア獣類の一目に位置づけられた。さらにその後複数の系統解析により、現生群の中では偶蹄目(鯨類も含む)に最も近縁であることが明らかになっている。奇蹄類と偶蹄類で真有蹄類(Euungulata) を成す。
絶滅した滑距目のマクラウケニアと南蹄目のトクソドンは、化石に残った分子情報による系統解析から、奇蹄目と姉妹群を成すことが明らかになっている。奇蹄目、南蹄目、滑距目などを纏めて「汎奇蹄類」(Panperissodactyla)とする分類が提唱されている。
系統的位置
- 北方真獣類の中での系統的位置
- 有蹄類の中での系統的位置
分類
- 哺乳綱 Mammalia
- 獣亜綱 Theria
- 真獣下綱 Eutheria
- ローラシア獣上目 Laurasiatheria
- 奇蹄目 Perissodactyla
- ローラシア獣上目 Laurasiatheria
- 真獣下綱 Eutheria
- 獣亜綱 Theria
奇蹄目の現生群はウマ科、サイ科そしてバク科の3科からなり、サイ科とバク科は有角類としてまとめられる。
絶滅群を含めた系統関係はあいまいであり、一例としてブロントテリウム科をウマ形亜目に含める、カリコテリウム科(鉤足類)の類縁と見なして月獣下目Selenidaに含める、独立した「ティタノテリウム型亜目」とするなど複数の説があり、分類は安定していない。カリコテリウム科(鉤足類)も有角類とともに「バク型亜目」に含める説がある。
以下の分類は、遠藤秀紀 (2002) に従う(†は絶滅群)。
- ウマ形亜目 Hippomorpha
- ウマ上科 Equoidea
- ウマ科 Equidae
- †パレオテリウム科 Palaeotheriidae
- †ブロントテリウム上科 Brontotherioidae(「ティタノテリウム型亜目」に分類する説もある)
- †ブロントテリウム科 Brontotheriidae(ラムドテリウム科を分ける説もある)
- ウマ上科 Equoidea
- †鉤足亜目 Ancylopoda(「バク型亜目」の鉤足下目とする説もある)
- †エオモロプス科 Eomoropidae
- †カリコテリウム科 Chalicotheriidae
- 有角亜目 Ceratomorpha(「バク型亜目」の有角下目とする説もある)
- バク上科 Tapiroidea
- †イセクトロプス科 Isectolophidae(有角類に含めない説もある)
- †ヘラレテス科 Helaletidae
- †ロフィアレテス科 Lophialetidae(有角類に含めない説もある)
- †デペレテラ科 Deperetellidae
- †ロフィオドン科 Lophiodontidae(鉤足類とする説もある)
- バク科 Tapiridae
- サイ上科 Rhinocerotoidea
- †ヒラコドン科 Hyracodontidae
- †アミノドン科 Amynodontidae
- サイ科 Rhinocerotidae
- バク上科 Tapiroidea
生態
有角亜目は繁殖期を除くと常時単独で行動しているが、ウマ形亜目はグループで生息している。
形態特徴
脚指の先端は蹄(ひづめ)で覆われており、指先のみを地面につけ、人間などのかかとに相当する部分は地面につけない蹄行性の歩行をする。各脚の指の数は、ウマでは1本、サイでは3本、バクでは前脚が4本、後脚が3本となっている。これらの指のうち、中指を肢端の中心線が通っていることがこのグループを定義付ける特徴である。かつて奇蹄目の祖先は5本の指を持っていたが、草原を走るため体重を支える第3指が発達し、他の指は退化している。
それ以外の四肢の特徴としては、大腿骨の外側に、第三小転子と呼ばれる筋肉の付着点となる突起を持ち、また距骨上端が滑車型、下端が平面または窪み型となっている点である。この距骨は、鯨偶蹄類では上下端とも滑車状になっている。
いずれも草食性で、切歯と臼歯がよく発達しており、食べ物を噛み切り、すり潰すのに適している。切歯は大半のものが上下三対とも揃っており、効率よく植物を裁断する。この切歯と臼歯の間には大きな間隙があり、犬歯を持つものは、この部分に孤立した形で生えていることが多い。臼歯のうち大臼歯は祖先的なグループでは丘状歯(ブノドント)であるが、より進化したグループでは畝状歯(ロフォドント)となっている。また、進化したグループでは、最前列を除く小臼歯が大臼歯とほぼ同じ形となっている。また進化につれ、高歯冠化が著しい。
胃は単室であるが、イヌやネコ、ヒトなど草食への特殊化をしていない単純な形態のものとは異なり、食道へと繋がる入口(噴門部)付近に憩室を備え、発酵室の機能を備えつつある傾向を持つ。下部消化管に関しては、結腸が発達し、食物を発酵・分解するためのタンクとしての役割を果たしている。また、結腸ほどではないが盲腸が長く発達している(ウマで約1.2m)。胆嚢が無いことも草食に適している。しかしながら大腸を発酵タンクとする方法は、反芻が行えない、肛門に近いため面積の確保が難しいなどのデメリットが挙げられる。また単胃で容量不足のため頻繁に採食しなければならず、反芻獣に比べて捕食されるリスクが大きいとの指摘もある。
出典
参考文献
- Nishihara H, Hasegawa M, Okada N (2006). “Pegasoferae, an unexpected mammalian clade revealed by tracking ancient retroposon insertions”. Proc Natl Acad Sci U S A 103 (26): 9929-34. doi:10.1073/pnas.0603797103.
- コルバート, エドウィン・ハリス、モラレス, マイケル、ミンコフ, イーライ・C.『脊椎動物の進化(原著第5版)』田隅本生訳、築地書房、2004年、400,449-472頁。ISBN 4-8067-1295-7。
- 遠藤秀紀『哺乳類の進化』東京大学出版会、2002年、92-99,186-190,222-235頁。ISBN 978-4-13-060182-5。
- 冨田幸光『絶滅哺乳類図鑑』伊藤丙雄、岡本泰子、丸善、2002年、140-159頁。ISBN 4-621-04943-7。
- 冨田幸光『絶滅哺乳類図鑑』伊藤丙雄、岡本泰子(新版)、丸善、2011年、161-181頁。ISBN 978-4621082904。
- 長谷川政美『新図説 動物の起源と進化 書き換えられた系統樹』八坂書房、2011年、43-44頁。ISBN 978-4-89694-971-1。
関連項目
- ウマ形亜目
- 髁節目
外部リンク
- 『奇蹄類』 - コトバンク